


临床分离侵袭性感染念珠菌药物敏感性及耐药机制流行病学特征

张志华,药理学博士,主任医师,教授,硕士生导师。现任河北北方学院附属第一医院院长、党委副书记、第一临床医学院院长。主要从事肺部肿瘤及呼吸危重症学研究,担任河北省呼吸专科联盟常务理事、河北省慢阻肺联盟常委、河北省医学会呼吸分会常务委员、河北省医学会急诊医学分会副主任委员、河北省抗癌协会常务理事、张家口市抗癌协会副理事长、河北省医师协会理事、河北省突发公共卫生事件应急处理专家、张家口市委研究室顾问、张家口大健康产业发展顾问等。近五年来先后承担国家科技重大专项横向课题,河北省财政厅、卫计委等科研项目二十余项。先后发表学术论文30余篇,其中SCI收录7篇,荣获河北省科技进步三等奖1项、市厅级科研奖项若干。

张伟,主管技师,河北北方学院附属第一医院中心实验室。主要从事病原微生物检验工作,研究方向为病原体耐药机制及流行病学。兼任河北省微生物学会青年委员。主持市厅级课题3项,以第一作者或共同第一作者发表论文5篇,其中SCI 2篇。
![]()
文丨张伟 詹明华 王新生 李景琦 苏英鹏 吴晨怡 陈新飞 侯欣 周梦兰 张志华
【摘要】念珠菌是引起人类相关疾病的重要病原体,可引起循环系统、神经系统、泌尿系统感染,在血流感染病原谱中排在第四位。白念珠菌约占所有念珠菌中一半比例,在念珠菌前五位中占首位,其次依次为光滑念珠菌、近平滑念珠菌、热带念珠菌、克柔念珠菌。临床一线抗真菌药物从唑类转为棘白菌素类后,念珠菌耐药形势也出现了变化。本文综述了国内外常见念珠菌的致病、耐药及耐药机制等流行病学特征,为相关研究提供参考。
【关键词】念珠菌;耐药机制;唑类;棘白菌素类;ERG11基因;FKS1基因
近年来,随着基质辅助激光解吸电离飞行时间质谱(MALDI/TOF-MS)技术、ITS基因测序技术以及宏基因组测序技术(m-NGS)应用于临床,病原体鉴定准确率得到了提升,一些罕见的病原体也逐渐被识别。“超级真菌”—耳念珠菌的出现给治疗其感染带来较大压力[1, 2]。了解念珠菌病原谱及药物敏感性流行病学特征,对于经验性治疗相关疾病及制定防御政策意义重大。
念珠菌可以引起皮肤表面浅部感染,也可引起循环系统、神经系统深部感染[3]。唑类药物(如氟康唑)由于具有毒性低、抗真菌谱广、可口服等优点,早期被视为首选抗真菌药物[4]。在一线药物由唑类转向棘白菌素类的背景下,念珠菌感染病例在上升,且念珠菌对唑类和棘白菌素类耐药性在增加[3, 5, 6]。常见念珠菌病原谱为白念珠菌、近平滑念珠菌、光滑念珠菌、热带念珠菌、克柔念珠菌,其中白念珠菌占比近一半,非白念珠菌占另一半[4, 7, 8]。
念珠菌药敏试验常用Sensititre YeastOne法和CLSI推荐的微量肉汤稀释法检测[6, 9]。Sensititre YeastOne法与参考方法相比结果具有一致性,但检测棘白菌素类时有一定的偏差,应按照M38-A2手册的指导进行念珠菌药敏试验[9, 10]。Etest被证明是一种很好的、易于操作的替代方法,用于检测对阿尼芬净和米卡芬净的敏感性,但不太适合检测卡泊芬净的敏感性[11]。唑类作用靶位在细胞膜,棘白菌素类则抑制细胞壁成分1,3-βD葡聚糖[3]。现将念珠菌致病特点及耐药机制进行如下综述。
一、白念珠菌致病特征及耐药机制
白念珠菌是侵袭性念珠菌感染最常见的病原体,约占所有临床分离念珠菌中一半比例,是机会致病念珠菌,可引起老年患者、免疫力下降患者血流感染、腹腔感染、泌尿系统感染,其中以血流感染多见,感染患者主要出现在ICU[3, 12-14]。白念珠菌可形成酵母相和假菌丝相来适应不同的环境[15]。由酵母相转变成菌丝相可促使白念珠菌生物膜形成,增强侵袭能力[12, 15, 16]。生物膜产生后,白念珠菌感染致病率和死亡率均会升高[17]。虽经积极治疗,但仍有40%白念珠菌感染患者死亡[16]。
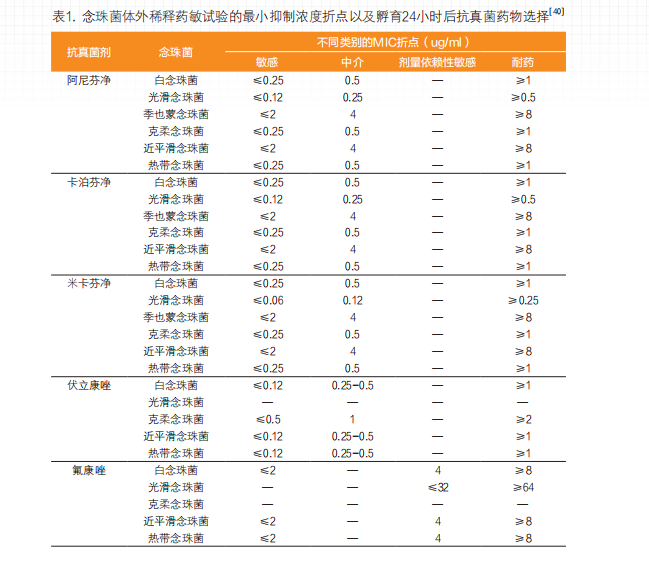
白念珠菌对氟康唑的耐药率为3.5%,流行病学届值(ECV)为0.12ug/ml[4]。瑞士25家医院一项长达10年的研究数据显示,白念珠菌对卡泊芬净、阿尼芬净和米卡芬净的ECV分别为0.12、0.06和0.03ug/ml[6]。新型抗真菌药物“瑞扎芬净”对白念珠菌的活性与阿尼芬净、米卡芬净和卡泊芬净相当,MIC50/90分别为0.03/0.06ug/ml[14]。CLSI临床分离念珠菌耐药折点(见表1)。
白念珠菌对唑类耐药涉及多种机制,包括甾醇成分改变导致ERG3失活、外排泵过表达、ERG11基因过表达、多倍体等[4]。体外检测白色念珠菌ERG11(CYP51)基因突变显示,Y132F、Y132H、K143R、F145L、S405F、G448E、G450E、G464S、Y132F和K143R、Y132F和F145L、D278N和G464S氨基酸替换可使氟康唑MIC至少增加2倍[18]。A114S,Y257H,E266D和V488I也是白念珠菌对唑类耐药的氨基酸替换位点[19]。Michelle L Villasmil等[20]发现了白念珠菌对唑类新的耐药机制,缺乏CaArv1基因的菌株对唑类呈高度敏感且毒性较低,抑制CaArv1基因的表达成为潜在治疗白念珠菌的新方法。人们常常关注ERG11(CYP51)基因与唑类药物耐药相关性,然而YongQin Wu等研究表明ERG11基因在白念珠菌的菌丝伸长、氧化应激适应和毒力方面起着重要作用,白念珠菌ERG11基因缺失可降低菌株菌丝生成能力,增加菌株对H2O2的敏感性,毒性降低,容易被巨噬细胞杀灭[21]。FKS1基因中的突变热点区2中R1361G氨基酸替换可导致白念珠菌对三种棘白菌素全耐药[6]。白念珠菌耐药机制模式图参考2021年发表的文献(见图1)。
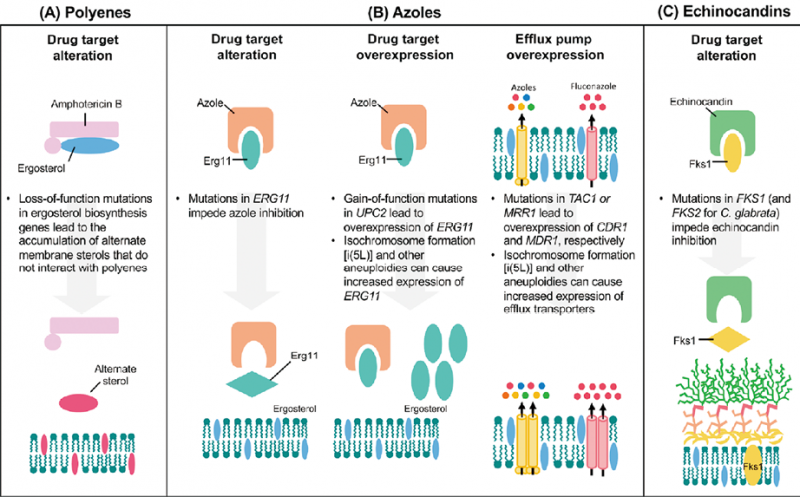
图1. 抗真菌耐药性的分子机制[41]
注:(A)对多烯的抗性主要是通过麦角甾醇生物合成基因的功能缺失突变而导致目标麦角甾醇的耗竭来介导的。这导致产生替代性甾醇,这些甾醇不能有效地与多烯相互作用,因此不能从真菌细胞膜中提取。
(B)对唑类药物的耐药性可能是通过替代唑类靶点Erg11而产生的,这导致对羊毛甾醇脱甲基酶的药物结合亲和力降低(左图)。药物靶点的过度表达可以通过转录激活剂UPC2中的功能获得突变,或通过非整倍体的形成,直接增加ERG11的拷贝数(中间图)。通过激活特定转录因子(白念珠菌中的TAC1和金黄色念珠菌中的TAC1以及光滑念珠菌中的PDR1)的突变,ABC转运蛋白(黄色)包括Cdr1和Cdr2的上调也可获得唑耐药性。此外,通过激活转录因子MRR1中的突变,MF转运体(粉红色)Mdr1的过度表达可导致唑类耐药性(右图)。外排泵也可以通过非整倍体形成而过度表达。
(C)棘白菌素耐药性主要涉及编码药物靶点(1,3)-β-D-葡聚糖合酶催化亚单位的FKS基因突变。对于白色念珠菌和耳道假丝酵母菌,FKS1中发生了赋予棘白菌素耐药性的突变,而对于光滑假丝酵母菌,FKS1及其同源FKS2中都发生了突变。
二、光滑念珠菌致病特征及耐药机制
光滑念珠菌是仅次于白念珠菌的第二大侵袭性念珠菌病原体,是由光滑念珠菌、尼瓦里氏假丝酵母菌和布拉卡伦斯假丝酵母菌组成的复合体[22-24]。SENTRY 2016-2018收集的1904株念珠菌中,光滑念珠菌占19.6%(373株)[14]。2003至2016年14年间光滑念珠菌在英国分离率没有增加,棘白菌素耐药非常罕见[25]。在使用棘白菌素治疗3-4周后,光滑念珠菌出现耐药,但是在没有使用棘白菌素的患者体内也检测出耐药的光滑念珠菌,证明耐药分离株可能在患者间存在传播[26]。
2016-2018年SENTRY的所有光滑念珠菌分离株中,8.6%表现出氟康唑耐药表型,有7.0%和12.8%的分离株分别对泊沙康唑和伏立康唑为非敏感[14]。在美国,光滑念珠菌对氟康唑的耐药率为7.8%,ECV为ug/ml[4]。米卡芬净、卡泊芬净、阿尼芬净对光滑念珠菌的MIC50/90分别为0.015/0.0μg/mL、0.03/0.06μg/mL、0.06/0.12μg/mL[14]。两性霉素B和唑类药物对光滑念珠菌的抗菌活性高[23]。2015-2018年英国棘白菌素耐药的光滑念珠菌仅占0.55%[25]。瑞扎芬净对光滑念珠菌的MIC值为0.06ug/ml,ECV为0.12ug/ml[14]。
光滑念珠菌耐药机制涉及外源性甾醇摄取、多倍体、ATP结合外排泵过表达、ERG11基因过表达,耐药机制见图2[4]。CDR1和CDR2过表达是光滑念珠菌常见的耐药原因之一,此外,ERG11基因中I166S氨基酸替换可导致光滑念珠菌对唑类耐药[19]。PFALLER M A等[14]研究表明,光滑念珠菌对棘白菌素类耐药由突变FKS2 HS1 S663P,FKS2 HS1 F659_del,FKS2 HS1 Y657_del/F658Y以及FKS1 HS1 S629P导致。英国2003-2016年14年间仅发现15株棘白菌素耐药光滑念珠菌,FKS1基因S629P和FKS2基因S663P氨基酸替换是导致耐药的主要原因[25]。ARASTEHFAR A等[27]研究表明,光滑念珠菌FKS2基因中S663P和F659缺失是导致棘白菌素耐药的原因。A. Kritikos等[6]也发现了FKS2基因的S663P突变可导致光滑念珠菌对三种棘白菌素全部耐药。
三、近平滑念珠菌致病特征及耐药机制
近平滑念珠菌是第三大侵袭性感染念珠菌病原体,由近平滑念珠菌、似平滑念珠菌、拟平滑念珠菌组成复合体,它的生物学特性与白念珠菌明显不同[24, 28]。SENTRY 2016-2018收集的1904株念珠菌中,近平滑念珠菌占17.3%(329株),拉丁美洲分离率最高,为20.2%,亚太地区分离率最低,为12.2%[14]。
近平滑念珠菌的耐药率在不同国家及地区见差异较大。WHALEY S G[4]等报道近平滑念珠菌对氟康唑的耐药率为3.4%,ECV为0.5ug/ml,但SENTRY 2016-2018统计近平滑念珠菌对氟康唑的耐药率为12.5%,主要来自欧洲医学中心[14]。土耳其在血液中分离的225株近平滑念珠菌中有27%对氟康唑耐药[27]。瑞扎芬净对近平滑念珠菌的MIC90值为2ug/ml,与米卡芬净(MIC90,1μg/mL;100.0% S)和阿尼芬净(MIC90,2μg/mL;93.9% S)相近,但比卡泊芬净(MIC90,0.5μg/mL;100.0% S)高4倍,ECV为≤4ug/ml[14]。
近平滑念珠菌耐药机制涉及外排泵过表达、ERG11基因突变[4]。ERG11基因中,单点突变Y132F和双点突变Y132F+K143R是导致近平滑念珠菌对唑类耐药的主要原因,Y132F突变株的患者死亡率和生物膜产生能力显著高于非突变株[27]。
四、热带念珠菌致病特征及耐药机制
热带念珠菌是常见的机会致病菌之一,在热带和亚热带地区尤为流行[29]。SENTRY 2016-2018收集的1904株念珠菌中,热带念珠菌占10.3%(190株)[14]。热带念珠菌大部分来自呼吸科和ICU,主要分离自呼吸道分泌物样本[30]。
WHALEY S G等[4]研究表明热带念珠菌对氟康唑的耐药率为2.3%,与PFALLER M A[14]等统计数据2.6%一致,ECV为0.25ug/ml。在中国,热带念珠菌对唑类敏感率、剂量依赖性敏感率和耐药率分别为76.7%、10.5%和12.8%[31]。值得注意的是,热带念珠菌对唑类药物耐药率逐年升高[30]。热带念珠菌对棘白菌素类敏感率高达99%,瑞扎芬净对热带念珠菌的MIC值为0.03ug/ml,ECV为0.12ug/ml[14]。
热带念珠菌对唑类耐药机制涉及甾醇成分变化使ERG3失活、ERG11基因过表达、ERG11基因突变、MDR1基因过表达[4, 31-33]。唑类耐药组的ERG11基因和UPC2基因的mRNA表达水平显著高于唑类敏感组,且这两个基因表达量呈显著的正相关性[30]。ERG11基因中的K143R氨基酸替换可导致热带念珠菌对多种唑类药物耐药,叠加ABC转运蛋白上调作用效果则更加凸显[24]。Y132F和S154F也是热带念珠菌ERG11基因中导致唑类耐药的突变靶点[19, 32, 33]。A395T/W,A395T/W+C461T/Y,V125A、Y257H和G464S均可导致热带念珠菌对唑类耐药,其中A395T/W和A395T/W+C461T/Y是最常见的耐药突变,而C461T/Y对耐药似乎不起作用[31]。在FKS1中的HS1的S645P和F650S可导致热带念珠菌对棘白菌素耐药,但有些菌株FKS1基因没有发生突变,仍可导致棘白菌素类MIC值升高[14]。
五、克柔念珠菌致病特征及耐药机制
SENTRY 2016-2018收集的1904株念珠菌中,克柔念珠菌占4%(76株)[14]。中国2015-2017年间分离的念珠菌中,克柔念珠菌占1%[34]。
克柔念珠菌对氟康唑天然活性较低[4]。瑞扎芬净对克柔念珠菌的MIC值为0.03ug/ml,ECV为0.12ug/ml;此外,米卡芬净、阿尼芬净和卡泊芬净对克柔念珠菌的敏感率分别为100%、100%和98.7%;伏立康唑和泊沙康唑对克柔念珠菌的敏感率分别为96.1%和100%[14]。CHIFNET分离的克柔念珠菌对卡泊芬净、米卡芬净和阿尼芬净的敏感率分别为87.2%、97.4%和100%[34]。
克柔念珠菌耐药机制涉及ATP结合外排泵过表达、多倍体、天然与唑类亲和力低[4]。
六、耳念珠菌致病特征及耐药机制
耳念珠菌是一种新发现的病原体,自从2009年被首次报道后,已在南极洲以外的所有大陆国家被分离出来,由于鉴定错误、高耐药率、高致死率给临床治疗带来较大挑战[1, 35-38]。耳念珠菌可在所有类型的患者中引起疾病,对环境消毒的抵抗力强,容易在环境中和患者之间传播[36]。耳念珠菌可以粘附于塑料载体上形成生物膜,但能力远不如白念珠菌[39]。在不同培养条件下,耳念珠菌可形成卵圆形、细长形和假菌丝形,其毒力低于白念珠菌[5]。
中国尚未分离出耐药的耳念珠菌[5]。然而在印度10家医院一项长达8年的研究数据显示,耳念珠菌对唑类、棘白菌素类和两性霉素B的耐药率分别为90%、2%和8%,多重耐药率和多唑类耐药率分别为25%和13%[2]。棘白菌素类对耳念珠菌显示出良好的杀菌活性,瑞扎芬净的活性不劣于或优于传统的棘白菌素类[35]。硫酸铜可抑制耳念珠菌生长[5]。
ERG11基因中Y132F和K143R氨基酸替换以及FKS1基因中突变热点区1 S639F氨基酸替换可导致耳念珠菌对唑类和棘白菌素类耐药[2, 38]。突变株与野生株的退火温度差超过4℃,利用此特性可用PCR法将二者区分[38]。
小结:念珠菌是人类不可忽视的病原体,虽然非白念珠菌占比有所增加,白念珠菌仍然占据重要权重。越来越多的机构报道了长期的监测数据显示,念珠菌耐药形势不容乐观,尤其是耳念珠菌在印度等国家的高耐药率需引起足够的警惕,需持续监测该菌的动向,防止出现全球大流行。
念珠菌对唑类耐药机制涉及甾醇成分改变/ERG3基因失活、外源性甾醇摄取、外排泵过表达、天然氟康唑亲和力低、多倍体、ERG11基因过表达、ERG11基因突变等(见图2)。
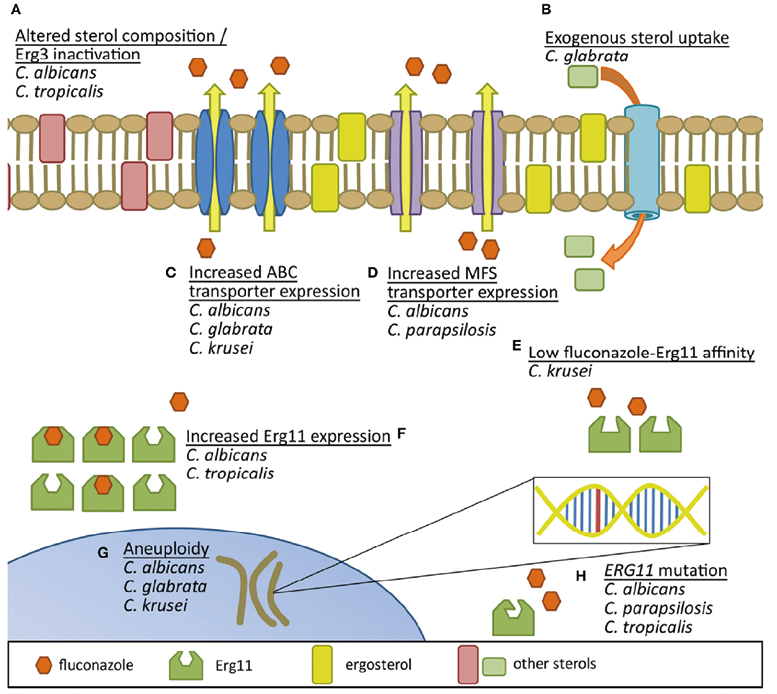
图2. 念珠菌对氟康唑耐药机制的比较[4]
注:(A)Erg3失活导致酵母膜中交替甾醇的利用。
(B)摄取外源性甾醇有助于避免氟康唑抑制内源性甾醇的产生。
(C)ATP结合盒外排泵和(D)主要促进剂超家族转运体产量的增加减少了唑类药物在细胞内
的积累。
(E)氟康唑固有的低亲和力结合物种特异性Erg11可能会降低氟康唑抑制该蛋白的潜力。
(F)Erg11蛋白表达的增加有助于克服唑的活性。
(G)非整倍体可能促进对唑暴露的遗传适应。
(H)ERG11突变也可导致氟康唑结合亲和力降低的蛋白质。
念珠菌对棘白菌素类耐药机制常见于FKS基因突变。不同种属间的念珠菌耐药机制有差异,随着人们对其认识的深入,有新的耐药机制逐渐被发现。在防控耐药病原体传播和耐药机制研究方面仍任重而道远。
参考文献
SPIVAK E S, HANSON K E. Candida auris: an Emerging Fungal Pathogen [J]. J Clin Microbiol, 2018, 56(2).
CHOWDHARY A, PRAKASH A, SHARMA C, et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009-17) in India: role of the ERG11 and FKS1 genes in azole and echinocandin resistance [J]. J Antimicrob Chemother, 2018, 73(4): 891-9.PRISTOV K E, GHANNOUM M A. Resistance of Candida to azoles and echinocandins worldwide [J]. Clin Microbiol Infect, 2019, 25(7): 792-8.
WHALEY S G, BERKOW E L, RYBAK J M, et al. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species [J]. Front Microbiol, 2016, 7: 2173.
WANG X, BING J, ZHENG Q, et al. The first isolate of Candida auris in China: clinical and biological aspects [J]. Emerg Microbes Infect, 2018, 7(1): 93.
KRITIKOS A, NEOFYTOS D, KHANNA N, et al. Accuracy of Sensititre YeastOne echinocandins epidemiological cut-off values for identification of FKS mutant Candida albicans and Candida glabrata: a ten year national survey of the Fungal Infection Network of Switzerland (FUNGINOS) [J]. Clin Microbiol Infect, 2018, 24(11): 1214.e1-.e4.
DÍAZ-GARCÍA J, MESQUIDA A, SÁNCHEZ-CARRILLO C, et al. Monitoring the Epidemiology and Antifungal Resistance of Yeasts Causing Fungemia in a Tertiary Care Hospital in Madrid, Spain: Any Relevant Changes in the Last 13 Years? [J]. Antimicrob Agents Chemother, 2021, 65(4).
BAI Y, ZHENG Z, LIU T, et al. Epidemiological Characteristics and Drug Resistance of Fungemia in General Hospitals from 2010 to 2019 [J]. Biomed Res Int, 2021, 2021: 2529171.
CHANG W, BAO F, WANG Z, et al. Comparison of the Sensititre YeastOne(®) and CLSI M38-A2 microdilution methods in determining the activity of nine antifungal agents against dermatophytes [J]. Mycoses, 2021, 64(7): 734-41.
CÓRDOBA S, ABIEGA C, AGORIO I, et al. [Usefulness of the Sensititre YeastOne® panel to detect Candida species resistant to antifungal drugs] [J]. Rev Argent Microbiol, 2021.
AIGNER M, ERBEZNIK T, GSCHWENTNER M, et al. Etest and Sensititre YeastOne Susceptibility Testing of Echinocandins against Candida Species from a Single Center in Austria [J]. Antimicrob Agents Chemother, 2017, 61(8).
LOHSE M B, GULATI M, JOHNSON A D, et al. Development and regulation of single- and multi-species Candida albicans biofilms [J]. Nat Rev Microbiol, 2018, 16(1): 19-31.
DADAR M, TIWARI R, KARTHIK K, et al. Candida albicans-Biology, molecular characterization, pathogenicity, and advances in diagnosis and control-An update [J]. Microb Pathog, 2018, 117: 128-38.
PFALLER M A, CARVALHAES C, MESSER S A, et al. Activity of a Long-Acting Echinocandin, Rezafungin, and Comparator Antifungal Agents Tested against Contemporary Invasive Fungal Isolates (SENTRY Program, 2016 to 2018) [J]. Antimicrob Agents Chemother, 2020, 64(4).
ARKOWITZ R A, BASSILANA M. Recent advances in understanding Candida albicans hyphal growth [J]. F1000Res, 2019, 8.
CHEN H, ZHOU X, REN B, et al. The regulation of hyphae growth in Candida albican[J]. Virulence, 2020, 11(1): 337-48.
PEREIRA R, DOS SANTOS FonTENELLE R O, DE BRITO E H S, et al. Biofilm of Candida albicans: formation, regulation and resistance [J]. J Appl Microbiol, 2021, 131(1): 11-22.
WARRILOW A G, NISHIMOTO A T, PARKER J E, et al. The Evolution of Azole Resistance in Candida albicans Sterol 14α-Demethylase (CYP51) through Incremental Amino Acid Substitutions [J]. Antimicrob Agents Chemother, 2019, 63(5).
TEO J Q, LEE S J, TAN A L, et al. Molecular mechanisms of azole resistance in Candida bloodstream isolates [J]. BMC Infect Dis, 2019, 19(1): 63.
VILLASMIL M L, BARBOSA A D, CUNNINGHAM J L, et al. An Erg11 lanosterol 14-α-demethylase-Arv1 complex is required for Candida albicans virulence [J]. PLoS One, 2020, 15(7): e0235746.
WU Y, WU M, WANG Y, et al. ERG11 couples oxidative stress adaptation, hyphal elongation and virulence in Candida albicans [J]. FEMS Yeast Res, 2018, 18(7).
LOTFALI E, FATTAHI A, SAYYAHFAR S, et al. A Review on Molecular Mechanisms of Antifungal Resistance in Candida glabrata: Update and Recent Advances [J]. Microb Drug Resist, 2021, 27(10): 1371-88.
HERNANDO-ORTIZ A, MATEO E, ORTEGA-RIVEROS M, et al. Caenorhabditis elegans as a Model System To Assess Candida glabrata, Candida nivariensis, and Candida bracarensis Virulence and Antifungal Efficacy [J]. Antimicrob Agents Chemother, 2020, 64(10).
XISTO M I, CARAMALHO R D, ROCHA D A, et al. Pan-azole-resistant Candida tropicalis carrying homozygous erg11 mutations at position K143R: a new emerging superbug? [J]. J Antimicrob Chemother, 2017, 72(4): 988-92.
FRASER M, BORMAN A M, THORN R, et al. Resistance to echinocandin antifungal agents in the United Kingdom in clinical isolates of Candida glabrata: Fifteen years of interpretation and assessment [J]. Med Mycol, 2020, 58(2): 219-26.
ARENDRUP M C, PATTERSON T F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment [J]. J Infect Dis, 2017, 216(suppl_3): S445-s51.
ARASTEHFAR A, DANESHNIA F, HILMIOĞLU-POLAT S, et al. First Report of Candidemia Clonal Outbreak Caused by Emerging Fluconazole-Resistant Candida parapsilosis Isolates Harboring Y132F and/or Y132F+K143R in Turkey [J]. Antimicrob Agents Chemother, 2020, 64(10).
TÓTH R, NOSEK J, MORA-MonTES H M, et al. Candida parapsilosis: from Genes to the Bedside [J]. Clin Microbiol Rev, 2019, 32(2).
XU J. Is Natural Population of Candida tropicalis Sexual, Parasexual, and/or Asexual? [J]. Front Cell Infect Microbiol, 2021, 11: 751676.
WANG D, AN N, YANG Y, et al. Candida tropicalis distribution and drug resistance is correlated with ERG11 and UPC2 expression [J]. Antimicrob Resist Infect Control, 2021, 10(1): 54.
FAN X, XIAO M, ZHANG D, et al. Molecular mechanisms of azole resistance in Candida tropicalis isolates causing invasive candidiasis in China [J]. Clin Microbiol Infect, 2019, 25(7): 885-91.
JIN L, CAO Z, WANG Q, et al. MDR1 overexpression combined with ERG11 mutations induce high-level fluconazole resistance in Candida tropicalis clinical isolates [J]. BMC Infect Dis, 2018, 18(1): 162.
JIANG C, DONG D, YU B, et al. Mechanisms of azole resistance in 52 clinical isolates of Candida tropicalis in China [J]. J Antimicrob Chemother, 2013, 68(4): 778-85.
XIAO M, CHEN S C, KONG F, et al. Distribution and Antifungal Susceptibility of Candida Species Causing Candidemia in China: An Update From the CHIF-NET Study [J]. J Infect Dis, 2020, 221(Suppl 2): S139-s47.
KOVÁCS R, TÓTH Z, LOCKE J B, et al. Comparison of In Vitro Killing Activity of Rezafungin, Anidulafungin, Caspofungin, and Micafungin against Four Candida auris Clades in RPMI-1640 in the Absence and Presence of Human Serum [J]. Microorganisms, 2021, 9(4).
SARIS K, MEIS J F, VOSS A. Candida auris [J]. Curr Opin Infect Dis, 2018, 31(4): 334-40.
SIKORA A, ZAHRA F. Candida Auris [M]. StatPearls. Treasure Island (FL); StatPearls Publishing Copyright© 2022, StatPearls Publishing LLC. 2022.
HOU X, LEE A, JIMÉNEZ-ORTIGOSA C, et al. Rapid Detection of ERG11-Associated Azole Resistance and FKS-Associated Echinocandin Resistance in Candida auris [J]. Antimicrob Agents Chemother, 2019, 63(1).
JEFFERY-SMITH A, TAORI S K, SCHELENZ S, et al. Candida auris: a Review of the Literature [J]. Clin Microbiol Rev, 2018, 31(1).
BARBARA D. ALEXANDER M, MHSGARY W,ET AL. Performance Standards for Antifungal Susceptibility Testing of Yeasts [J]. CLINICAL AND LABORATORY STANDARDS INSTITUTE, 2017, M60(1st Ed).
LEE Y, PUUMALA E, ROBBINS N, et al. Antifungal Drug Resistance: Molecular Mechanisms in Candida albicans and Beyond [J]. Chem Rev, 2021, 121(6): 3390-411.
基金资助项目:张家口市重点研发项目(2121098D和2121064D);
河北北方学院自然科学研究计划项目(XJ2021019)
作者单位:075000 河北张家口,河北北方学院附属第一医院中心实验室(张伟 詹明华 王新生 李景琦 苏英鹏 吴晨怡 张志华);
100730 北京,中国医学科学院北京协和医院检验科,疑难重症及罕见病国家重点实验室(张伟 陈新飞 侯欣 周梦兰)
通讯作者:张志华


