


布鲁氏菌病细胞免疫研究与临床应用前景

姜海,博士,研究员,硕士生导师,中国疾病预防控制中心传染病预防控制所布鲁氏菌病室副主任。中国地方病协会布鲁氏菌病专业委员会主任委员,生态健康与人类生态专业委员会委员,中国研究型医院学会生物标志物专业委员会委员。主要从事布鲁氏菌病快速诊断标志物及分子流行病研究。主持和参与了国家重点研发、科技重大专项和国家自然基金等多项国家级科研项目。在《Emerging Infectious Disease》,《Emerging Microbes & Infections》,《Infectious Diseases of Poverty》,《Frontiers in Microbiology》等SCI收录杂志上发表学术论文近30余篇。2020年荣获“全国卫生健康系统新冠肺炎疫情防控工作先进个人”称号。2021年,荣获中华预防医学科技奖三等奖。

韩阔,中国疾病预防控制中心传染病预防控制所2021级病原生物学专业硕士研究生,主要研究方向为布鲁氏菌病的细胞免疫学研究。
【摘要】布鲁氏菌病由布鲁氏菌感染引起,是世界范围内最常见的人畜共患病之一。布鲁氏菌是一种兼性细胞内感染细菌,能够通过其毒力因子调节宿主细胞信号通路并逃避宿主细胞免疫反应。巨噬细胞、树突状细胞(DC)以及T淋巴细胞可以通过各种不同途径参与机体抗布鲁氏菌的过程。不同宿主感染布鲁氏菌后诱导的细胞免疫反应及其细胞因子不尽相同。本文简要介绍了布鲁氏菌毒力因子介导的免疫逃逸机制,并在此基础上主要介绍了巨噬细胞、树突状细胞以及T淋巴细胞不同亚群参与的机体抗布鲁氏菌过程,还重点介绍了人类感染布鲁氏菌诱导不同T细胞亚群反应并分泌不同细胞因子。为了解布鲁氏菌病的发病机制,治疗以及疫苗研制提供了重要的依据。
【关键词】布鲁氏菌;巨噬细胞;树突状细胞;Th1细胞;Th2细胞;细胞因子
布鲁氏菌属于兼性细胞内革兰氏阴性菌,能够引起人畜共患病布鲁氏菌病。布鲁氏菌病在世界上许多国家流行,是全球最常见的人畜共患疾病之一,每年有超过50万人类新发病例[1]。布鲁氏菌主要通过吸入、皮肤擦伤、摄入或黏膜侵入机体[2]。布鲁氏菌通过其毒力因子脂多糖(LPS)、T4SS分泌系统、BvrR/BvrS系统和外膜蛋白,这些可以与宿主细胞表面相互作用,形成早期和晚期BCV(Brucella Containing Vacuole,布氏小体)以及细菌繁殖时与内质网(ER)的相互作用,介导其逃避免疫系统攻击。
布鲁氏菌最重要的特征是在吞噬细胞和非吞噬细胞内存活和繁殖并且可以改变免疫细胞(巨噬细胞、树突状细胞、T淋巴细胞等)介导的免疫反应,进而导致淋巴结、肝脏和脾脏肿大,该菌还具有抵抗吞噬细胞杀菌作用的能力,并且可以阻止抗原特异性T细胞对其识别,从而形成有利于其生存和繁殖的微环境,导致宿主慢性感染[3-6]。
机体对抗布鲁氏菌主要以细胞免疫应答为主,主要包括巨噬细胞、DCs等抗原提呈细胞及T淋巴细胞的活化,不同的T淋巴细胞亚群参与布鲁氏菌病的不同阶段。CD4+T淋巴细胞亚群,如Th1和Th2细胞,参与细菌感染,Th1通过分泌IFN-γ和IL-2等细胞因子在细菌感染中起重要作用。它可以消除细胞内细菌感染。Th1通常在感染的早期阶段发挥作用,并在宿主对病原体的抵抗力中起关键作用,特别是对于细胞内细菌感染。Th2细胞在感染中的主要功能是诱导体液免疫。Th17细胞与Th1细胞之间具有调节关系,Th17细胞的生长和发育受到IFN-γ和IL-4的负调节。成熟的Th17细胞可以抵抗IFN-γ和IL-4的抑制。Th17细胞与Th1和Th2炎症细胞系不同,能够弥补Th1/Th2作用机制的不足[7,8]。
本文主要讨论了主要免疫细胞(巨噬细胞、树突状细胞、T淋巴细胞各亚群)及不同物种在感染布鲁氏菌时的不同反应以及各免疫细胞所分泌的不同细胞因子。通过这些途径寻求布鲁氏菌病诊断、治疗及疫苗研制策略。
一、布鲁氏菌通过其毒力因子逃避宿主免疫反应
布鲁氏菌不产生经典的毒力因子:外毒素、外酶、质粒等。主要的毒力因子是:脂多糖(LPS),T4SS分泌系统,BvrR/BvrS系统和外膜蛋白等。布鲁氏菌的可以通过LPS的不同组分抵抗先天性免疫防御[9, 10, 11];通过VirB编码的T4SS分泌系统的形成并通过其释放的效应因子(BtpA)来调节宿主细胞信号通路并抑制宿主细胞免疫反应,此外T4SS还参与布鲁氏菌细胞内循环,促进机体宿主细胞形成布氏小体(BCV)并促进宿主细胞间感染[12, 13, 14, 15];BvrS/BvrR系统可通过调控布鲁氏菌外膜蛋白、LPS、VirB操纵子的表达对细菌逃避宿主细胞免疫识别以及感染新的细胞产生重要的影响[16];布鲁氏菌的外膜蛋白可以保护这些细菌免受宿主补体和其他抗菌肽的侵害,除此之外外膜蛋白还被证明可以介导布鲁氏菌和宿主细胞之间的直接相互作用,进而促进布鲁氏菌逃避宿主免疫反应[17, 18, 19, 20]。
二、布鲁氏菌介导的细胞免疫应答
布鲁氏菌感染(以牛种菌为例)后启动先天性和适应性免疫,TLR可参与识别布鲁氏菌,TLR2、TLR4和TLR9是参与识别牛种菌最重要TLR,在感染的早期阶段,不同的TLRs激活不同信号通路介导的巨噬细胞(Mø)和树突状细胞(DC)分泌IL-12和TNF-α。而布鲁氏菌产生的Toll/白细胞介素-1受体(TIR)可参与降低TLR对感染的依赖性反应。此外,IL-10中性粒细胞(Ne)增加CD35、CD11b和IL-8的表达,降低CD62表达,这会导致组织损伤;感染早期阶段的巨噬细胞IL-10增加,这会下调IFN-γ、IL-12和TNF-α的产生,导致布鲁氏菌可以在巨噬细胞中存活。而在感染的后期阶段,活化的巨噬细胞的杀菌活性主要是由T辅助1型(Th1)CD4细胞产生的IFN-γ诱导的活性氮中间体(RNIs)和活性氧中间体(ROIs);感染后DC的活化除了上调MHC II类(MHC II)和共刺激分子CD40、CD80和CD86的表达外,还会分泌IL-12和TNF-α。巨噬细胞和DC产生TNF-α和IL-12,刺激CD4和CD8T淋巴细胞分泌IFN-γ并发挥细胞毒性;Th1细胞反应包括CD4T细胞产生的IFN-γ,Th1CD4T细胞也增加CD28的表达并降低CTLA-4的表达,而在Th2CD4T细胞中的情况则相反。Th2反应的特征主要是CD4T细胞分泌IL-4和IL-5,这有利于慢性感染。CD8细胞毒性(Tc1)T细胞通过细胞溶解活性杀死受感染的宿主细胞(图1)[21]。
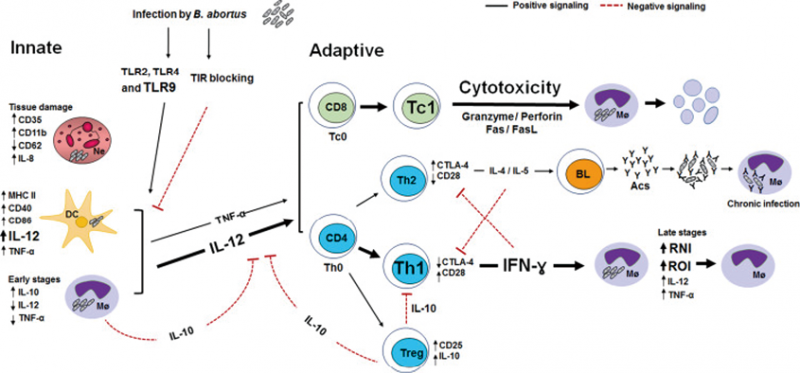
图1. 感染牛种菌后不同先天性和适应性免疫系统细胞的关键机制
1. 巨噬细胞对布鲁氏菌感染的反应:布鲁氏菌可在吞噬细胞、非吞噬细胞或其他细胞(如上皮样细胞,HeLa)中存活和繁殖,但这种细菌的主要靶细胞是巨噬细胞、树突状细胞和滋养层细胞。布鲁氏菌侵入巨噬细胞后,在最初的4h内,80%~90%的细菌被杀死,存活下来的布鲁氏菌会驻留在巨噬细胞囊泡中形成布氏小体并诱导VirBT4SS表达,进而通过一系列反应促进布鲁氏菌细胞内循环、细胞间的感染和T4SS效应蛋白的表达,从而调节宿主细胞信号通路并逃避宿主细胞免疫反应[15]。据报道,光滑型LPS对布鲁氏菌在巨噬细胞中生存具有免疫调节作用,它可以与巨噬细胞表面的MHCⅡ结合,形成MHCⅡ-LPS复合物进而干扰巨噬细胞以MHCⅡ的方式提呈抗原,从而削弱巨噬细胞活化抗原特异性CD4+T细胞的能力[22]。此外,布鲁氏菌LPS可通过巨噬细胞表面的脂筏进入细胞并与TNF-α相互作用来抑制宿主细胞凋亡,从而促进布鲁氏菌在宿主细胞中的生存与复制[23]。另外,布鲁氏菌外膜蛋白也可以诱导机体产生免疫应答,例如:Omp16与Omp19能够跟巨噬细胞TLR2作用,激活细胞释放IFN-α、IL-6、IL-10与IL-12等促炎因子,此外Omp19可下调IFN-γ的释放,进而抑制人类单核细胞的抗原提呈功能[24]。此外,布鲁氏菌感染会使单核细胞失调,并通过上调LC3依赖的自噬过程抑制巨噬细胞分化为经典活化的巨噬细胞(M1巨噬细胞)或旁路活化的巨噬细胞(M2巨噬细胞),从而抑制促炎和抗炎因子的产生并干扰机体的免疫反应[25]。
2. 树突状细胞对布鲁氏菌感染的反应:树突状细胞(DC)是最早遇到抗原的细胞之一,这些细胞能够触发先天细胞和T细胞反应,并且是最有效的抗原呈递细胞。布鲁氏菌可以扩散并到达不同的淋巴组织,并能够感染不同的DC亚群。在布鲁氏菌感染早期,CD4、CD25T细胞产生的IL-10在急性期具有调节作用,可减少炎症和组织损伤,这种免疫反应可以保护细菌免于被消灭,此外早期IL-10的产生对于进化为慢性布鲁氏菌病至关重要[26]。布鲁氏菌LPS是否阻碍DC免疫反应存在一些争议,一些实验报告了光滑布鲁氏菌菌株对DC成熟或激活具有抑制作用,而感染粗糙菌株后,可对DC成熟具有激活作用[27]。大量研究表明,布鲁氏菌可被DC亚群的TLRs识别并激活细胞内途径产生细胞因子。如:当GM-DC被纯布鲁氏菌Omp19刺激时,会阻断TLR2,细胞因子的产生会受到抑制[28];热杀灭的布鲁氏菌(HKBa)可以在脾脏DC中与TLR2相互作用,并通过p38发出信号,导致吞噬作用,进而产生TNF-α导致吞噬体融合;然后TLR9与晚期内体中的细菌DNA相互作用,为IL-12的产生发出信号[29]。
此外,布鲁氏菌的外膜蛋白在DC成熟与抗原提呈方面发挥着重要的作用,例如:外膜蛋白Omp25能够抑制TNF-α的分泌,从而抑制受布鲁氏菌感染的DCs的成熟并且还可以抑制提呈抗原给初始性T细胞的功能,并且可抑制IL-12的分泌[30];而Omp19能够促进DCs的成熟与T细胞向Th1免疫应答[31]。除此之外研究表明,双突变布鲁氏菌(znBAZ)在被巨噬细胞吞噬后会释放IL-12和IL-18诱导早期NK细胞活化,激活的NK细胞会释放颗粒酶B和IFN-γ,颗粒酶B诱导感染细胞死亡,而IFN-γ诱导DC激活并摄取细菌抗原或吞噬受感染的宿主细胞,在摄取细菌抗原后,DC迁移到淋巴结能够引发CD8T细胞激活分化,并迁移到感染部位以提供保护[32]。
3. T淋巴细胞对布鲁氏菌感染的反应:布鲁氏菌主要通过胃肠道和呼吸道黏膜进入体内并诱发黏膜免疫反应,IFN-γ、IL-17以及Th1淋巴细胞介导的特异性免疫反应在布鲁氏菌感染黏膜的免疫反应中发挥着十分重要的作用。据报道,在用卵白蛋白和布鲁氏菌U-Omp19作为佐剂免疫的小鼠中,Th1、Th17和CD8+T淋巴细胞的抗卵白蛋白反应在肠道黏膜系统中增加,并且IFN-γ和IL-17会大量产生[33];此外,IFN-γ和IL-17还会影响肺部布鲁氏菌的控制[34]。布鲁氏菌对宿主的入侵主要诱导由Th1淋巴细胞介导的特异性免疫反应,该特异性免疫反应能够产生促炎细胞因IL-1β,IL-6,TNF-α,最为重要的是IFN-γ,这些促炎细胞因子会激活巨噬细胞以增加其杀菌能力,并激活杀死感染细胞的细胞毒性T淋巴细胞(CTL)从而发挥保护作用[35]。此外Th1细胞可以通过增加IL-2的分泌来对布鲁氏菌感染起免疫应答。
Th2细胞参与布鲁氏菌病的慢性感染,但在急性感染中,大多数血清Th2细胞因子的水平也会升高。Th2细胞主要通过分泌IL-4、IL-5和IL-13参与对抗布鲁氏菌感染。Th17细胞可以在布鲁氏菌感染后分化和增殖。Th17细胞由IL-6,IL-21和TGF-β诱导,IL-17A是Th17细胞的主要效应分子。Th17细胞可能参与细胞免疫对抗布鲁氏菌感染,但其具体机制需要进一步研究[36]。
三、布鲁氏菌介导人类的免疫应答
在人群中,布鲁氏菌主要通过胃肠道和呼吸道粘膜进入体内进而导致人类感染布鲁氏菌病,并表现出流感样症状、典型的波状热和骨关节炎等慢性感染[37]。细胞介导的免疫反应对保护人类免受布鲁氏菌感染方面有着十分重要的作用。在细胞介导的免疫过程中,抗原呈递细胞和T淋巴细胞帮助宿主抵抗或清除布鲁氏菌感染[38]。
T细胞免疫反应参与布鲁氏菌感染的全过程,但不同阶段(急性、慢性和恢复期)的患者会诱发不同的T细胞亚群免疫反应。相关研究表明:Th1、Th17和Treg细胞免疫在急性期占主导地位,而Th2、Th17和Treg细胞免疫在慢性期占主导地位[39]。Th1细胞的比例在人布鲁氏菌病患者中增加,是布鲁氏菌感染急性期细胞免疫的主要类型,IFN-γ是Th1细胞产生的关键细胞因子,在急性感染患者中,IL-2、IFN-γ和IL-10水平显著增加,Th1细胞通过产生IFN-γ激活巨噬细胞和细胞毒性T淋巴细胞[40]。另外,IL-12通过诱导IFN-γ的产生综合了对布鲁氏菌的先天和适应性免疫。IL-12和IFN-γ作为Th1型细胞因子,反过来诱导更多的Th1细胞极化,并在预防布鲁氏菌感染中发挥主要作用[41]。此外,Th1细胞还可以通过增加IL-2的分泌来对布鲁氏菌感染起免疫应答[39]。Th1细胞能够分泌TNF-α,对病原体和炎症的生化反应起着至关重要的作用。研究发现布鲁氏菌病患者和健康受试者之间的IFN-γ/TNF-α比率存在显著差异,重度组和非重度组之间也存在显著差异,因此这个比率也被认为可以作为疾病严重程度的有用指标[42]。
Th2细胞主要分泌的细胞因子是IL-4、IL-5和IL-13。据报道,Th2细胞在慢性布鲁氏菌病患者外周血中有较高的表达率,并且明显高于急性布鲁氏菌病患者[39]。IL-4的表达在慢性患者中表达率较高,显著高于急性患者。慢性患者IL-4表达的增加与Th2的变化一致,这说明IL-4与慢性布鲁氏菌感染有关[43]。除此之外,慢性布病患者中IL-5和IL-13的表达显著高于急性期,恢复期和健康人群,这表明IL-5和IL-13都参与Th2细胞介导的慢性布鲁氏菌感染[39]。有研究表明:IL-25(也称为IL-17E)能够增加Th2相关细胞因子(包括IL-4)的分泌,刺激体液反应[44],但最近的一项研究发现:IL-25在感染组与非感染组中没有明显差异[45]。
Th17细胞是与感染相关的重要免疫细胞,可参与细胞免疫对抗布鲁氏菌感染,并且可以在布鲁氏菌感染后增殖分化。据报道,急性布鲁氏菌病患者外周血中Th17细胞的百分率明显升高,治疗后下降[46],另外急性、慢性患者间的细胞比例没有显著差异,但高于健康对照组[39]。Th17细胞及其效应分子(IL-17、IL-17A和IL-17F),可以参与对布鲁氏菌感染的细胞免疫[47]。研究表明:急性和慢性患者Th17细胞的效应分子IL-17A和IL-17F的表达水平高于健康对照组[39]。此外还有研究表明:布鲁氏菌病患者血清IL-17水平高于健康对照组[48]。
四、总结与前景
布鲁氏菌通过其毒力因子降低抗原提呈细胞的吞噬能力与抗原提呈能力,延缓细胞成熟,抑制细胞凋亡,减少相关细胞因子的释放,干扰特异性与非特异性免疫反应。机体对布鲁氏菌的抵抗主要以细胞免疫应答包括巨噬细胞、DCs等抗原提呈细胞及不同T淋巴细胞亚群活化为主。布鲁氏菌通过与免疫细胞相互作用可引起各种细胞因子的释放,各类细胞因子既可以激活免疫细胞的杀菌作用并诱导受感染细胞凋亡,也可以促进免疫细胞的抗原提呈作用及共刺激分子的表达并引起T淋巴细胞分化。
疫苗是用于控制布鲁氏菌病的重要组成部分,但目前没有安全有效的疫苗可用于预防人类布鲁氏菌病,并且布鲁氏菌病是一种难以治疗的细胞内细菌感染疾病。通过对布鲁氏菌与免疫细胞之间相互作用以及它们所分泌的细胞因子的研究,可以更好地理解布鲁氏菌病的状况、疗效和预后的生化或免疫指标。相信在未来,我们不仅可以将这些用于诊断,还用于开发疫苗。![]()
参考文献
Georgios P, et al. The new global map of human brucellosis[J]. Lancet Infect Dis, 2006.
Franco MP, Mulder M, Gilman RH, et al. 2007. Human brucellosis. Lancet Infect Dis. 7(12): 775–786.
Roop RM, Gaines J M, Anderson E S, et al. Survival of the fittest: how Brucella strains adapt to their intracellular niche in the host[J]. Med Microbiol Immunol, 2009, 198(4): 221-238.
Barrionuevo P, Cassataro J, Delpino M V, et al. Brucella abortus inhibits major histocompatibility complex class II expression and antigen processing through interleukin-6 secretion via Toll-like receptor 2.[J]. Infect Immunity, 2008, 76(1): 250.
Ficht, Thomas, A, et al. Pathogenesis and Immunobiology of Brucellosis Review of Brucella-Host Interactions[J].American Journal of Pathology: Official Publication of the American Association of Pathologists, 2015.
Perkins S D, Smither S J, Atkins HS. Towards a vaccine for humans[J]. 2010, 34(3): 379-394.
Pacholczyk R, Kraj P, Ignatowicz L. Peptide specificity of thymic selection of CD4+CD25+ T Cells[J]. J Immunol, 2002, 168(2): 613.
Sakaguchi S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses.[J]. An Re Immunol, 2004, 22(22): 531-562.
Smith J A. Brucella Lipopolysaccharide and pathogenicity: The core of the matter.[J]. Virulence, 2018(1).
Barquero-Calvo E, Mora-Cartín R, Arce-Gorvel V, et al. Brucella abortus Induces the premature death of human neutrophils through the action of its lipopolysaccharide[J]. PLoS Pathogens, 2015, 11(5): e1004853.
Jimenezd BMP, Terraza A, Gross A, et al. Different responses of macrophages to smooth and rough brucella spp.: relationship to virulence[J]. Infect Immunity, 2004, 72(4): 2429.
Ke Y, Wang Y, Li W, et al. Type IV secretion system of Brucella spp. and its effectors. Front Cell Infect Microbiol. 2015; 5:72.
Herrou J, Crosson S. Molecular structure of the brucella abortus metalloprotein ricA, a rab2-binding virulence effector[J]. Biochemistry, 2013, 52(50): 9020-9028.
Alaidarous M, Ve T, Casey L W, et al. Mechanism of bacterial interference with TLR4 signaling by brucella Toll/Interleukin-1 receptor domain-containing protein TcpB[J]. J Biol Chem, 2014, 289(2): 654-668.
Jiao H. The VirB System Plays a Crucial Role in Brucella Intracellular Infection[J]. International Journal of Molecular Sciences, 2021, 22.
Altamirano-Silva P, Cordero-Serrano M, Méndez-Montoya J, et al. Intracellular passage triggers a molecular response in brucella abortus that increases its infectiousness[J]. Infect immunity: IAI.00004-21.
Degos C, Hysenaj L, Gonzalez Spinoza G, et al. Omp25-dependent engagement of SLAMF1 by brucella abortus in dendritic cells limits acute inflammation and favours bacterial persistence in vivo[J]. Cell Microbiol, 2020, 22(4).
Ma Q L, Liu A C, Ma X J, et al. Brucella outer membrane protein Omp25 induces microglial cells in vitro to secrete inflammatory cytokines and inhibit apoptosis[J]. Internat J Clini Exp Med, 2015, 8(10): 17530-17535.
LN Velásquez, Milillo M A, Delpino M V, et al. Brucella abortus down-regulates MHC class II by the IL-6-dependent inhibition of CIITA through the downmodulation of IFN regulatory factor-1 (IRF-1)[J]. J Leuk Biol, 2017, 101(3).
Pasquevich K A, Carabajal M V, Guaimas F F, et al. Omp19 enables brucella abortus to evade the antimicrobial activity from host's proteolytic defense system data_sheet_1. pdf [J]. Front Immunol, 2019, 10.
Dorneles E, Teixeira-Carvalho A, MSS Araújo, et al. Immune response triggered by Brucella abortus following infection or vaccination[J]. Vaccine, 2015, 33(31): 3659.
Chen F, Ding X, Ding Y, et al. Proinflammatory caspase-2-me-diated macrophage cell death induced by a rough attenuatedBrucella suis strain[J].Infect Immun, 2011, 79(6): 2460-2469. DOI: 10.1128/IAI.00050-11.
Lapaque N, Moriyon I, Moreno Eet al. 2005. Brucella lipopolysaccharide acts as a virulence factor. Curr Opin Microbiol. 8(1): 60–66.
Pasquevich KA, Estein SM, García Samartino C, et al. Immu-nization with recombinant Brucellaspecies outer membrane pro-tein Omp16or Omp19in adjuvant induces specific CD4+andCD8+T cells as well as systemic and oral protection againstBrucella abortus infection[J].Infect Immun, 2009, 77(1): 436-445. DOI: 10.1128/IAI.01151-08.
Yang W, Li Y, Li H, et al. Brucella dysregulates monocytes and inhibits macrophage polarization through LC3-Dependent autophagy[J]. Front Immunol, 2017, 8.
Xavier M N, Winter M G, Spees A M, et al. CD4+ T cell-derived IL-10 promotes brucella abortus persistence via modulation of macrophage function[J]. PLOS Pathogens, 9, 6(2013-6-20), 2013, 9(6): e1003454.
Eric Daniel AC, Flores-Romo L, Sharon W, et al. Dendritic cells and Brucella spp. interaction: the sentinel host and the stealthy pathogen.[J]. Springer Netherlands, 2020(1).
Zhang K, Wang H, Guo F, et al. OMP31 of Brucella melitensis 16M impairs the apoptosis of macrophages triggered by TNF-α[J]. Exp Therap Med, 2016, 12(4): 2783.
Zhang CY, Bai N, Zhang ZH, et al. TLR2 signaling subpathways regulate TLR9 signaling for the effective induction of IL-12 upon stimulation by heat-killed Brucella abortus. Cell Mol Immunol. 2012; 9(4): 324–333. doi: 10.1038/cmi.2012.11.
Billard E, Dornand J, Gross A . Brucella suis prevents human dendritic cell maturation and antigen presentation through regulation of tumor necrosis factor alpha secretion.[J]. Infect Immunity, 2007, 75(10): 4980-4989.
Zwerdling A, Delpino M V , Barrionuevo P , et al. Brucella lipoproteins mimic dendritic cell maturation induced by Brucella abortus[J]. Micr Infect, 2008, 10(12-13):1346-1354.
Bhagyaraj E, Wang H, Yang X, et al. Mucosal vaccination primes NK cell-dependent development of CD8+ T cells against pulmonary brucella infection. Front Immunol. 2021 Jul 7;12:697953. doi: 10.3389/fimmu.2021.697953. PMID: 34305935; PMCID: PMC8293993.
Andrés Esteban Ibañez, Coria L M, Marianela Verónica C, et al. A bacterial protease inhibitor protects antigens delivered in oral vaccines from digestion while triggering specific mucosal immune responses[J]. J Cont Re, 2015.
Hanot Mambres D, Machelart A, Potemberg G, et al. Identification of immune effectors essential to the control of primary and secondary intranasal infection with brucella melitensis in mice[J]. J Immunol, 2016, 196(9): 3780-3793.
Byndloss M X, Tsolis R M. Brucella spp. Virulence factors and immunity.[J]. An Rev An Biosciences, 2016, 4(1):111.
Tang Yunxia, et al. “Serum levels of seven general cytokines in acute brucellosis before and after treatment.” Infection and drug resistance vol. 14 5501-5510. 18 Dec. 2021, doi:10.2147/IDR.S341331
R López-Santiago AB, Sánchez-Argáez LGD, Alba-Núñez, et al. Immune response to mucosal brucella infection[J]. Front Immunol, 2019, 10: 1759-.
Skendros P, Pappas G, Boura P. Cell-mediated immunity in human brucellosis[J]. Micr Infect, 2011, 13(2):134-142.
Zheng R, Xie S, Zhang Q, et al. Circulating Th1, Th2, Th17, Treg, and PD-1 levels in patients with brucellosis. J Immunol Res. 2019 Aug 6; 2019: 3783209. doi: 10.1155/2019/3783209. PMID: 31467933; PMCID: PMC6701318.
N, Jain-Gupta A, et al. Immunotherapeutics to prevent the replication of Brucella in a treatment failure mouse model[J]. Vaccine, 2014, 32(8):918-923.
Eskandari-Nasab M, et al. Levels of interleukin-(IL)-12p40 are markedly increased in brucellosis among patients with specificIL-12BGenotypes[J]. Sc J Immunol, 2013, 78(1): 85-91.
Xu G, Zhang P, Dang R, et al. Dynamic changes of th1 cytokines and the clinical significance of the IFN-γ /TNF-α ratio in acute brucellosis[J]. Med Inflammat, 2019, 2019:1-10.
Akbulut H, Celik I, Akbulut A. Cytokine levels in patients with brucellosis and their relations with the treatment. In J Med Microbiol. 2007;25(4):387–390. doi: 10.4103/0255-0857.37345.
Zeng W P . 'All things considered': transcriptional regulation of T helper type 2 cell differentiation from precursor to effector activation.[J]. Immunology, 2013, 140(1):31-8.
Gheitasi R, et al. “evaluation of Th2 and Th17 Immunity-Related Factors as Indicators of Brucellosis.” Frontiers in cellular and infection microbiology vol. 11 786994. 7 Jan. 2022, doi:10.3389/fcimb.2021.786994
Masoomeh S, Amitis R, Alia M. et al. Interlukine-17 and TGF-β levels in patients with acute brucellosis before and after treatment[J]. Tur J Med En, 2016, 46: 1348-1352.
Zhu, Yongzhang et al. “Key immunity characteristics of diverse stages of brucellosis in rural population from Inner Mongolia, China.” Infectious diseases of poverty vol. 11,1 63. 4 Jun. 2022, doi:10.1186/s40249-022-00989-7
Lin ZQ, Lin GY, He WW, et al. IL-6 and INF-gamma levels in patients with brucellosis in severe epidemic region, Xinjiang, China. Infect Dis Poverty. 2020; 9(1): 47. doi: 10.1186/s40249-020-00666-7.


